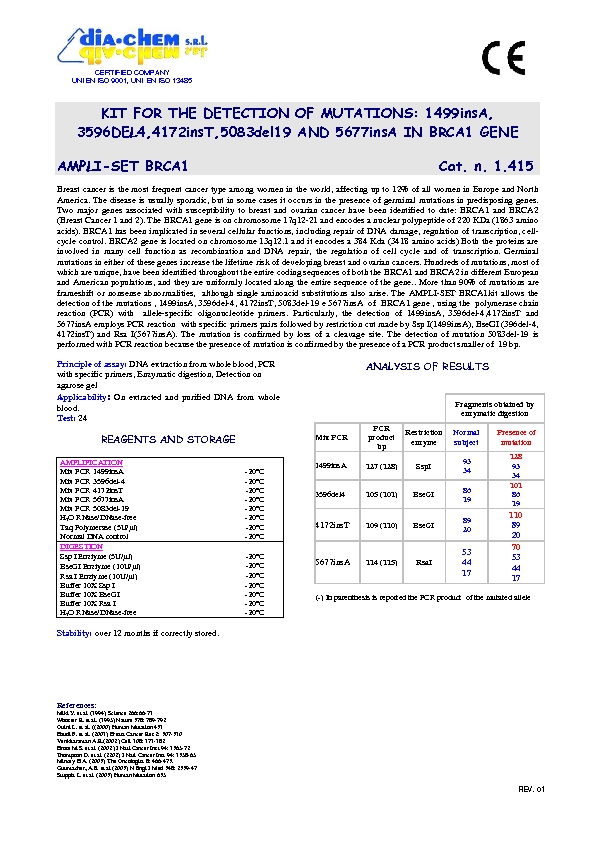
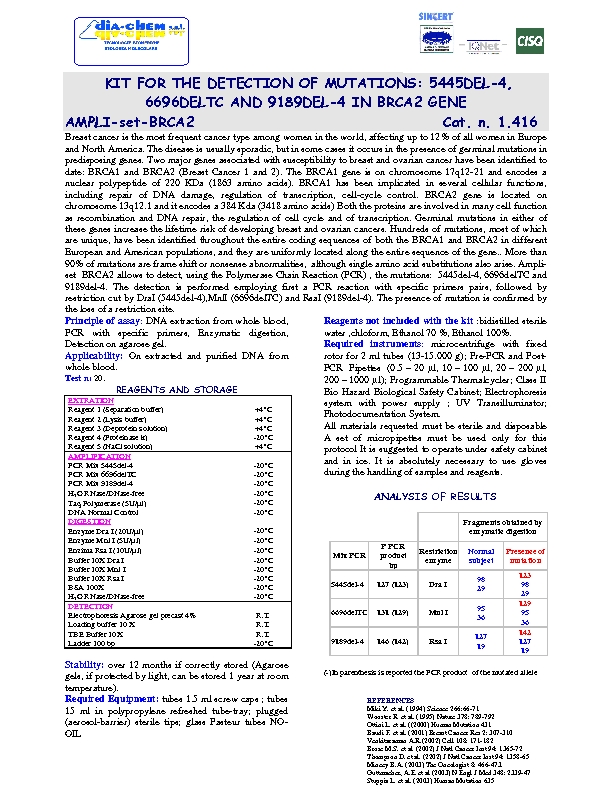
Il tumore, oggi, può essere considerato una patologia a componente genetica caratterizzata da una crescita cellulare incontrollata. Le cellule del nostro corpo ricevono dei segnali che indicano loro quando crescere e moltiplicarsi e quando tale crescita deve arrestarsi. Nel tumore tali cellule, a causa di alterazioni del proprio patrimonio genetico, non rispondono ai segnali di controllo e crescono e si moltiplicano irregolarmente dif- fondendosi in diverse parti del corpo. L'evento che determina l’alterazione della funzione dei geni viene definito mutazione. Quando un gene subisce una mutazione per cause biologiche, chimiche, fisiche, le in- formazioni che arriveranno alla cellula saranno improprie per le funzioni a cui è deputata. La maggior parte dei tumori sono sporadici, cioè le alterazioni del DNA si sviluppano casualmente a livello delle cellule so- matiche, cioè quelle cellule che costituiscono ogni organo ed apparato del nostro organismo. Queste muta- zioni si originano nel DNA di un ristretto gruppo di cellule e determineranno l’errore genetico che si perpe- tuerà nelle discendenti di quelle cellule, le quali accumulandosi in un determinato organo si sostituiranno inizialmente al tessuto sano per poi diffondersi in altri organi vicini o a distanza (metastasi).
La metilazione del DNA è un processo post-replicativo. Tra le modificazioni riguardanti il DNA, la metila- zione è fondamentalmente decisa durante lo sviluppo. La metilazione del DNA è quindi uno dei meccani- smi correlati con il differenziamento cellulare, tramite l’inibizione dell’espressione genica a livello trascri- zionale. E’ essenziale per il normale sviluppo dei mammiferi, è associata all’imprinting genomico, all’inat- tivazione trascrizionale del cromosoma X, all’invecchiamento ed ha un ruolo nello sviluppo di eventi pato- logici, come la tumorogenesi. La metilazione del DNA è una modificazione epigenetica post-sintetica che, con il trasferimento di un gruppo metilico dalla S-adenosylmetionina all’atomo di C in posizione 5 dell’a- nello della Citosina, introduce la 5mC come nuova base nel DNA.
Le CpG islands contengono dinucleotidi CpG con una frequenza matematicamente predeterminata. Le CpG islands sono circa 30.000 generalmente localizzate all'estremità 5’ della regione promotore di geni hou- sekeeping, qualche volta sovrapponendosi alla regione codificante per una estensione variabile (di solito il primo esone). La frequenza con cui i dinucleotidi CpG sono presenti nel genoma è inferiore alle aspettative, fuorché per le regioni CpG islands. Questo è il risultato di un meccanismo evolutivo legato alla presenza di una spontanea attività deaminasica nel nucleo. Questa reazione enzimatica trasforma la Citosina metilata in timina e quella non metilata in Uracile. Controlli seguenti e meccanismi di riparazione riconoscono l‘Uracile come base estranea del DNA e perciò la sostituiscono, mentre questa sostituzione non avviene per la Timina, base comune nel DNA. La maggior parte delle CpG non sono metilate nella cellula normale indi- pendentemente dallo stato trascrizionale del gene, mentre durante lo sviluppo del tumore, le CpGs al di fuo- ri delle CpG islands divengono ipometilate, e le CpG islands nella regione del promotore dei geni oncosop- pressori divengono ipermetilate. Questa ipermetilazione è associata alla condensazione della cromatina ed alla perdita di trascrizione. I dati più recenti indicano che gli eventi epigenetici e genetici interagiscono tra loro per aiutare lo sviluppo progressivo del tumore.